Os primeiros organismos que existiram eram unicelulares. O modo como estes se tornaram multicelulares é um grande passo evolutivoe existe consequentemente um grande debate sobre isso. Ao contrário de outros grandes passos na evolução, julga-se que a multicelularidade terá surgido várias vezes independentemente umas das outras - plantas, animais e fungos tiveram vias evolutivas independentes, e vários outros clados evoluíram multicelularidade.
Criacionismo e design inteligente
Design Inteligente
Se você estivesse caminhando e encontrasse uma pedra no caminho? Não seria de causar surpresa, uma vez que a pedra é uma estrutura bruta formada de compostos naturais que estão naquele lugar há milhares de anos!
Mas se ao caminhar você encontrasse um relógio? Você saberia que aquele objeto não foi formado naquele lugar, mas sim por alguém, uma vez que devido sua complexidade mecânica não poderia ser algo natural daquele ambiente.
Esse é o princípio do Design Inteligente, o fato de existir vida e da forma como ela é, complexa, não poderia ser creditada ao acaso!
Projeto inteligente ou design inteligente é a tradução do termo inglês intelligent design, corrente de pensamento que busca contestar as ideias evolucionistas em relação ao surgimento da vida na Terra e à seleção natural.
A base do ideal dessa corrente “científica” é a afirmação de que a diversidade biológica não se deu evolutivamente, mas sim por interferência ou condução de uma inteligência superior, não reportando essa ação a Deus ou a seres extraterrestres.
O Projeto Inteligente ganhou visibilidade graças a pressão de comunidades religiosas nos Estados Unidos, que judicialmente conseguiram inserir os conteúdos do Projeto Inteligente nas escolas. Em alguns casos os conteúdos evolucionistas foram simplesmente suprimidos do currículo.
A tentativa de dificultar o ensino de conceitos científicos, que não atendam aos ensinamentos religiosos, não é novidade nos Estados Unidos. Em 1925, no estado do Tennessee, foi promulgada uma lei que estabelecia que o professor que ensinasse qualquer teoria contrária à bíblica seria preso e quanto às escolas, as que ensinassem teorias evolutivas, teriam suas verbas estaduais cortadas. O julgamento da causa da lei gerou o título “Julgamento do Macaco”, como caracterização da polêmica gerada.
Para alguns cientistas, a apresentação do Projeto Inteligente seria apenas uma reformulação da teoria religiosa criacionista, que busca se contrapor às teorias científicas sobre o processo de evolução, principalmente às ideias de Darwin.
Como toda teoria, o Design Inteligente se apóia em três argumentos primários:
1. Complexidade Irredutível: Refere-se ao fato de a vida ser composta de partes interligadas que dependem umas das outras para que sejam úteis. A mudança em uma parte apenas por mutação, por exemplo, não poderia ser responsável pela eficiência de toda estrutura.
2. Complexidade Específica: Apresenta que seria impossível que padrões tão complexos, como os presentes nos seres vivos, tenham se desenvolvido através de processos do acaso.
3. Princípio Antrópico: Acredita que a existência e desenvolvimento da vida na Terra requerem que tantas variáveis estejam perfeitamente harmonizadas, que seria impossível que todas as variáveis chegassem a ser como são apenas pelo acaso. Se nosso planeta fosse um pouco mais próximo do Sol, as condições para existência de vida seriam inviáveis.
No Brasil, a tradução do livro A Caixa Preta de Darwin - O Desafio da Bioquimica à Teoria da Evolução, pela editora Jorge Zahar Editor, do autor Behe, constitui um importante meio de informar aos interessados sobre as ideias do Desing Inteligente.
Criacionismo
O Criacionismo é a teoria ou crença religiosa na qual a humanidade, toda a vida na Terra e no Universo são frutos da criação de um ser sobrenatural.
Essa teoria parte do pressuposto de que essas criações não estariam sujeitas a evoluções ou transformações. Ela se opõe as teorias evolucionistas, baseadas principalmente nos estudos de Charles Darwin.
O criacionismo é um discurso baseado em tradições escritas que buscam explicações literais para narrativas mítico-religiosas presentes nas escrituras de diferentes religiões.
Assim, as religiões do mundo irão construir sua própria teoria criacionista.
Na civilização grega, foi com os Titãs que se deu a origem do homem, moldado em barro, como na narrativa bíblica, na qual o homem teria sido criado após o céu e terra.
Vale destacar que três importantes religiões monoteístas possuem suas versões criacionistas e são amplamente aceitas pela população: o judaísmo, cristianismo e islamismo.
De modo geral, todas as civilizações buscam ou buscaram dar conta da origem do Homem e do Universo e cada uma possui uma versão sobre o tema.
Na maioria, a explicação é religiosa, a qual afirma que todos os seres vivos teriam sido criados por uma ou mais entidades (quase sempre antropomórficas).
Isso porque, o Criador faz sua criatura à sua imagem e semelhança.
Observe que nas crenças dessa natureza, a interferência divina se estende para além do ato de criação. O ser divino continua agindo no mundo mortal, como quando Deus inspira Noé a construir sua arca para fugir do dilúvio.
Criacionismo Científico e a oposição ao evolucionismo
Apesar do termo Criacionismo se aplicar a todas as explicações mítico-religiosas para origem da vida no Universo, este termo ficou mais conhecido a partir de sua oposição à ciência evolutiva do século XVIII.
Nesse momento, ele esteve exposto o contraste entre as teorias científicas, como a da evolução, e as explicações religiosas.
Portanto, a partir das explicações evolucionistas científicas, especialmente as de Charles Darwin, durante o século XIX, os argumentos criacionistas irão fazer suas revisões e reinterpretações.
Esse movimento fica mais claro nos Estados Unidos, onde o termo "criacionismo" torna-se popular a partir da década de 1920.
Nesse momento, ele passou a ser associado ao posicionamento fundamental do cristianismo para o tema da evolução humana, especialmente entre as igrejas protestantes naquele país.
Muitas vezes os criacionistas irão rejeitar as conclusões das pesquisas científicas. Contudo, alguns irão se apoiar no cientificismo para constituir argumentos que vão além do cunho religioso.
Isso porque incorporam atributos científicos e filosóficos para fundamentar as evidências de que houvera planejamento no que tange ao surgimento das espécies (Design Inteligente).
Importante destacar ainda que há uma posição criacionista que afirma a existência de um Criador sem identificá-lo, enquanto outras afirmam quem é o Criador.
Fósseis e a evolução
A vida multicelular é um dos grandes marcos da evolução biológica. Ela permitiu o surgimento de organismos complexos e diversificados, como plantas, animais e fungos. No entanto, o momento exato em que a vida multicelular surgiu na Terra ainda é um mistério. Uma nova descoberta, publicada na revista Science, pode ajudar a resolver esse mistério. Pesquisadores da Universidade de Oxford, no Reino Unido, encontraram fósseis de organismos multicelulares com 1,6 bilhão de anos de idade. Esses fósseis são minúsculos, com apenas alguns milímetros de diâmetro. Eles foram encontrados em rochas sedimentares da África do Sul. Os pesquisadores acreditam que eles representavam organismos semelhantes a algas, com células especializadas que desempenhavam funções diferentes. Essa descoberta é significativa porque sugere que a vida multicelular surgiu muito mais cedo do que se pensava anteriormente. A teoria mais aceita até então afirmava que a vida multicelular surgiu há cerca de 1,2 bilhão de anos. A descoberta também levanta novas questões sobre a evolução da vida multicelular. Por que a vida multicelular surgiu tão cedo? Quais foram as forças evolutivas que levaram ao surgimento de organismos complexos e diversificados? Essas são questões que os cientistas ainda estão tentando responder. No entanto, a descoberta desses fósseis minúsculos é um importante passo em direção a uma melhor compreensão da história da vida na Terra. Aqui estão alguns pontos importantes sobre a descoberta:
Os fósseis foram encontrados em rochas sedimentares da África do Sul.
Eles têm cerca de 1,6 bilhão de anos de idade.
Eles representam organismos semelhantes a algas, com células especializadas.
Essa descoberta sugere que a vida multicelular surgiu muito mais cedo do que se pensava anteriormente.
O que essa descoberta significa para nós? Essa descoberta é importante porque nos ajuda a entender melhor a história da vida na Terra. Ela sugere que a vida multicelular surgiu muito mais cedo do que se pensava anteriormente. Isso tem implicações importantes para nossa compreensão da evolução biológica. A descoberta também nos dá pistas sobre as forças evolutivas que levaram ao surgimento de organismos complexos e diversificados. Isso pode nos ajudar a entender melhor como a vida evoluiu na Terra e como ela pode evoluir no futuro.
As evidências da evolução
Existem três grandes tipos de explicação evolutiva para a explosão que conduziu à disparidade de Burgess. O primeiro é convencional e tem sido aceito em quase todas as discussões publicadas. Os dois últimos possuem pontos em comum e representam tendências recentes no pensamento evolutivo.
1) O primeiro Enchimento do Barril Ecológico:
Na teoria darwiniana convencional, os organismos propõem e o ambiente dispõe. Os organismos fornecem a matéria prima na forma de variação genética que se expressa através de diferenças morfológicas. Dentro de uma população, em qualquer tempo, essas variações são pequenas e não direcionadas. A modificação evolutiva é produzida por forças de seleção natural provenientes do ambiente externo (tanto condições físicas, como interações com outros organismos). Como os organismos proporcionam apenas a matéria-prima e como essa matéria-prima quase sempre tem sido considerada suficiente para todas as modificações que se processam de acordo com os ritmos darwinianos, caracteristicamente lentos, o ambiente transforma-se no motor que regula a velocidade e a extensão das alterações evolutivas. Então quando nos perguntamos que peculiaridade ambiental poderia ter engendrado a explosão Cambriana, uma resposta óbvia nos vem de imediato à mente: A explosão Cambriana foi o primeiro enchimento do barril ecológico da vida multicelular. Foi uma época de oportunidades inigualáveis. Quase todas as formas de vida podiam encontrar seu lugar ao sol. A vida estava se irradiando para os espaços vazios e poderia proliferar em progressão logarítmica, tal como uma célula bacteriana numa placa de ágar. Na efervescência desse período inigualável, a experimentação imperou pela primeira vez e última vez num mundo praticamente desprovido de competição.
“Competição menos severa” tornou-se a senha da interpretação. Whittington, escreveu:
“Havia provavelmente alimento e espaço abundantes nos diversificados ambientes que inicialmente foram sendo ocupados por esses novos animais, e a competição era menos severa do que nos períodos posteriores. Nessas circunstâncias, diversas combinações de características podem ter sido possíveis, enquanto estavam sendo desenvolvidas novas formas de captar os estímulos sensoriais dos arredores, de obter alimento, de fazer locomoção.... É assim que podem ter surgido estranhos animais que não se encaixam em nossas classificações dos quais alguns remanescentes podemos ver em Burgess Shale”.
2) Uma história direcional para sistemas genéticos
Na visão darwiniana tradicional, as morfologias possuem histórias que restringem seus futuros, embora o material genético não “envelheça”. Diferenças nas taxas e nos padrões de mudança constituem respostas de um substrato material inalterável (genes) às variações ambientais que reajustam as pressões da seleção natural. Mas talvez os sistemas genéticos realmente “envelheçam” no sentido de se tornarem “menos propensos a perderem uma grande reestruturação” (Valentine). Talvez os organismos modernos não possam gerar uma rápida sucessão de designs fundamentalmente novos, quaisquer que sejam as oportunidades ecológicas. A descoberta da fauna pré-cambriana de Ediacara, com forte possibilidade de que este primeiro conjunto de fósseis multicelulares talvez não tenha dado origem aos grupos modernos, sugere que todos os animais do cambriano, a despeito da disparidade de formas, podem ter divergido não muito antes a partir de um ancestral comum do final do Pré-cambriano. Se isto ocorreu – se eles ficaram separados apenas por um curto período de tempo – todos os animais cambrianos podem ter sido portadores de um mecanismo genético muito similar em virtude do seu tempo estritamente limitado de vida independente. “A resposta similar dos organismos de Burges pode refletir a homologia de um sistema genético em grande parte ainda conservado em comum e ainda bastante flexível, e não apenas a analogia da resposta a uma pressão externa comum. É claro que a vida teve necessidade do impulso externo da oportunidade ecológica, mas sua capacidade de resposta pode ter indicado uma herança genética comum, agora dissipada”.
3) Diversificação inicial e posterior fechamento como uma propriedade de sistemas.
Atu Kauffman, da Universidade da Pensilvânia, desenvolveu um modelo para demonstrar que o padrão de Burges, caracterizado por rápida disparidade máxima seguida pela posterior dizimação, é uma propriedade geral dos sistemas. Para explicar seu modelo, Kauffman considera metaforicamente, o palco da vida como uma paisagem complexa, com milhares de picos de alturas diferentes. Quanto mais elevado for o pico, maior o êxito dos organismos que estão sobre ele. Sobre os picos dessa paisagem estão espalhados ao acaso uns poucos organismos iniciais. Eles se multiplicarão e trocarão de posições. Os saltos grandes podem ser definidos como aqueles que levam um organismo para tão longe do seu antigo lar que o novo panorama não guarda nenhuma relação com o antigo. Os grandes saltos são extremamente arriscados mas proporcionam uma enorme recompensa para aqueles que obtêm sucesso. Se você aterrisa num pico mais alto do que aquele onde se encontrava, você prospera e se diversifica, se aterrisa num pico mais baixo ou num vale, você está liquidado. Kauffman demonstra que a probabilidade de êxito é no início bastante alta mas logo cai e chega a praticamente zero – tal como na história da vida. Se os saltos longos ocorrem com razoável frequência, todos os picos altos serão ocupados logo no início. Assim, os vitoriosos cavam trincheiras e a evolução produz sistemas desenvolvimentais tão ligados aos seus picos que, mesmo que surja uma nova oportunidade, as espécies não poderão mais mudar. Mesmo extinções ao acaso deixarão espaços inacessíveis a todos.
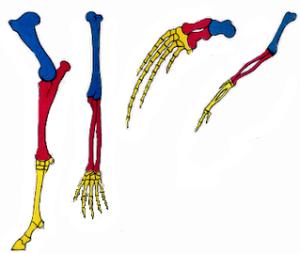
Pata dianteira de um equino, braço humano, nadadeira peitoral de um mamífero e asa de uma ave: homologias
No mundo científico, as hipóteses são elaboradas como respostas para determinadas perguntas acerca de um fenômeno específico. Quando uma hipótese é confirmada diversas vezes, por experimentações e/ou um conjunto de evidências, ela tem grandes chances de se tornar uma teoria.
Assim, a Teoria da Evolução reúne uma série de evidências e provas que a faz ser irrefutável até o presente momento:
A primeira evidência refere-se aos registros fósseis, sendo uma prova consistente de que nosso planeta já abrigou espécies diferentes das que existem hoje. Esses registros são uma forte evidência da evolução porque podem nos fornecer indícios de parentesco entre estes e os seres viventes atuais ao observarmos, em muitos casos, uma modificação contínua das espécies.
A adaptação, capacidade do ser vivo em se ajustar ao ambiente, pode ser outra evidência, uma vez que, por seleção natural, indivíduos portadores de determinadas características vantajosas - como a coloração parecida com a de seu substrato - possuem mais chances de sobreviver e transmitir a seus descendentes tais características. Assim, ao longo das gerações, determinadas características vão se modificando, tornando cada vez mais eficientes. Como exemplos de adaptação por seleção natural temos a camuflagem e o mimetismo.
As analogias e homologias também podem ser consideradas como provas da evolução baseadas em aspectos morfológicos e funcionais, uma vez que o estudo comparativo da anatomia dos organismos mostra a existência de um padrão fundamental similar na estrutura dos sistemas de órgãos.
Estruturas análogas desempenham a mesma função, mas possuem origens diferenciadas, como as asas de insetos e asas de aves. Estas, apesar de exercerem papéis semelhantes, não são derivadas das mesmas estruturas presentes em um ancestral comum exclusivo entre essas duas espécies. Assim, a adaptação evolutiva a modos de vida semelhantes leva organismos pouco aparentados a desenvolverem formas semelhantes, fenômeno este chamado de evolução convergente.
Homologia se refere a estruturas corporais ou órgãos que possuem origem embrionária semelhante, podendo desempenhar mesma função (nadadeira de uma baleia e nadadeira de um golfinho) ou funções diferentes, como as asas de um morcego e os braços de um humano, e nadadeiras peitorais de um golfinho e as asas de uma ave. Essa adaptação a modos de vida distintos é denominada evolução divergente.
Os órgãos vestigiais – estruturas pouco desenvolvidas e sem função expressiva no organismo, como o apêndice vermiforme e o cóccis - podem indicar que estes órgãos foram importantes em nossos ancestrais remotos e, por deixarem de ser vantajosos ao longo da evolução, regrediram durante tal processo. Estes órgãos podem, também, estar presentes em determinadas espécies e ausentes em outras, mesmo ambas existindo em um mesmo período.
Uma última evidência, a evidência molecular, nos mostra a semelhança na estrutura molecular de diversos organismos sendo que, quanto maior as semelhanças entre as sequências das bases nitrogenadas dos ácidos nucleicos ou quanto maior a semelhança entre as proteínas destas espécies, maior o parentesco e, portanto, a proximidade evolutiva entre as espécies.
As ideias de Lamarck
Lamarck defendia que a matéria gelatinosa inanimada podia espontaneamente gerar formas simples de vida, através da força dos fluidos ativos, a saber: água, terra, fogo e ar. A essa forma mais simples de vida Lamarck chamou de germe.
Esses germes podiam, por sua vez, gerar plantas ou animais por meio de progressões contínuas de uma forma para outra ao longo das gerações. As espécies se modificavam das formas mais simples em direção às formas mais complexas ao longo do tempo, buscando alcançar a perfeição da espécie. Assim, organismos que descendem de linhagens mais antigas seriam mais complexos apenas porque tiveram mais tempo para evoluir.
Lamarck não supôs que diferentes espécies descendem de ancestrais comuns, mas que são frutos de gerações espontâneas distintas. Cabe ressaltar dois fenômenos que, segundo Lamarck, contribuem para a biodiversidade encontrada no planeta:
o processo de geração espontânea dar-se-ia de forma continuada;
as linhagens produzidas podem progredir por caminhos evolutivos diversos, em função do ambiente que elas ocuparam durante a trajetória.
Lamarck não acreditava, então, em extinção de espécies. Para ele, espécies desapareceram ao longo da história porque evoluíram, se transformando em outras espécies diferentes. Vamos ver como se dá esse processo.
Lamarck também defendia que o uso contínuo de determinado órgão provocaria o seu desenvolvimento. Em contrapartida, um órgão em desuso poderia ser diminuído e até extinto. Essa explicação ficou conhecida como a primeira lei de Lamarck.
A ideia dessa lei é que fatores ambientais podiam alterar os movimentos dos fluidos internos dos animais, abrindo novas passagens entre as células e criando novos órgãos.
A base dessa lei foi a comparação de órgãos de recém-nascidos com órgãos dos mesmos animais na fase adulta. Todas as modificações encontradas foram atribuídas aos movimentos dos quatro elementos.
O exemplo mais famoso de Lamarck para explicar o desenvolvimento de órgãos muito usado foi o do pescoço das girafas. Segundo ele, o hábito de as girafas esticarem o pescoço para comer as folhas das copas das árvores fez com que o membro ficasse cada vez mais comprido ao longo das gerações, até chegar ao tamanho que conhecemos hoje.
Figura 2: Girafa.
Nenhum dos exemplos dados por Lamarck, seja para a perda de órgãos em desuso, seja para o desenvolvimento de órgãos muito usados, ficou tão famoso quanto esse do aumento do pescoço da girafa.
Entre os outros exemplos dados por ele, podemos citar a perda da visão das toupeiras e a perda da capacidade de voar das aves mantidas em cativeiro.
Figura 3: Toupeiras. Difícil de ver, não? Elas ficam enterradas!
Lamarck defendia que todas as mudanças estruturais sofridas por um indivíduo seriam transmitidas para seus descendentes. Isso explicaria tanto o alongamento do pescoço das girafas quanto a cegueira das toupeiras. A hereditariedade das características adquiridas ou perdidas ficou conhecida como a segunda lei de Lamarck.
Cabe ressaltar que Lamarck reconhecia que algumas mudanças só seriam possíveis ao longo de várias gerações, ou seja, se considerássemos períodos de tempo muito longos.
Lamarck também estudou sementes e plantas ao longo de sua vida. Uma de suas observações foi a de que sementes de uma mesma espécie cultivadas em altitudes diferentes podem gerar plantas com características estruturais bem diversas.
Suas observações na área da Botânica mostraram que o clima e a disponibilidade de recursos exerciam forte influência na constituição física dos organismos, corroborando a explicação dada por Lamarck para a enorme biodiversidade encontrada no planeta.
Apesar de as teorias de Lamarck terem sido desacreditadas ao longo da história em razão das pesquisas e descobertas de outros cientistas, sua importância para o surgimento da ideia de evolução não foi diminuída. A estátua em sua homenagem que fica na entrada dos jardins das plantas em Paris, traz a seguinte inscrição: "Fundador da doutrina da evolução".
Figura 4: Estátua de Lamarck na entrada dos Jardins da Plantas, Paris.
Teoria de Darwin
Charles Darwin era um naturalista britânico que propôs a teoria da evolução biológica por seleção natural.
Darwin definiu evolução como "descender com modificações", a ideia de que as espécies mudam ao longo do tempo, dão origem a novas espécies e compartilham um ancestral comum.
O mecanismo que Darwin propôs para evolução é a seleção natural. Em razão dos recursos limitados, organismos com características hereditárias que favoreçam a sobrevivência e a reprodução tendem a deixar mais descendentes do que os demais, o que faz com que essas características aumentem em frequência ao longo das gerações.
A seleção natural faz com que as populações se tornem adaptadas, ou cada vez mais bem integradas a seus ambientes ao longo do tempo. A seleção natural depende do ambiente e requer a existência de variações genéticas em um grupo.
Evolução
Darwin propôs que as espécies podem mudar ao longo do tempo, que novas espécies surgem de espécies pré-existentes, e que todas compartilham um ancestral comum. Nesse modelo, cada espécie tem um conjunto único de diferenças herdáveis (genéticas) comparadas ao ancestral comum, que se acumularam gradualmente ao longo do tempo. Eventos repetidos de diferenciação, nos quais espécies novas divergem a partir de seu ancestral comum, produzem uma "árvore" multinível que conecta todos os seres vivos.
Darwin se refere a esse processo no qual os organismos mudam suas características herdáveis através das gerações como "descendência com modificação". Atualmente, nós chamamos de evolução. Os rascunhos de Darwin vistos acima ilustram suas ideias, mostrando como uma espécie pode se diferenciar em duas com o passar do tempo, e como esse processo pode se repetir muitas e muitas vezes na "árvore genealógica" de um grupo de espécies relacionadas.
Seleção Natural
Darwin, é importante notar, não propôs apenas que os organismos evoluíam. Se esse tivesse sido o começo e o fim de sua teoria, ele não estaria em tantos livros didáticos como está hoje! Ao invés, Darwin também propôs um mecanismo para a evolução: seleção natural. Esse mecanismo era elegante e lógico, e explicava como populações podiam evoluir (passar por descendência com modificação) de tal maneira que se tornassem melhor adaptadas aos seus ambientes ao longo do tempo.
O conceito de seleção natural de Darwin foi baseado em algumas observações importantes:
Características são geralmente herdáveis. Em seres vivos, muitas características são herdadas, ou passadas dos pais para os filhos. (Darwin sabia que era esse o caso, apesar de não saber que as características eram herdadas através de genes.)
Um diagrama com um texto que diz: os progenitores transmitem características hereditárias para seus descendentes. À esquerda, uma borboleta azul-escuro e uma borboleta azul-claro cruzam para produzir descendentes com asas em vários tons de azul. À direita, uma borboleta vermelho-escuro e uma borboleta vermelho-claro cruzam para produzir descendentes com asas em vários tons de vermelho.
Nem toda a prole é capaz de sobreviver. Os organismos são capazes de produzir uma prole maior do que o ambiente é capaz de suportar. Por isso, há competição por recursos limitados em cada geração.
Um diagrama com uma caixa que diz: recursos limitados. Setas apontam para fora da caixa em direção a balões com os textos: falta de alimentos, falta de habitat e falta de parceiros. O texto abaixo disso diz: …nem todos os indivíduos sobreviverão e se reproduzirão. Há uma imagem de um grupo de 16 borboletas com asas em vários tons de azul e de vermelho. Um balão contém o texto glup! que vem de 4 das borboletas.
A prole varia quanto às características herdáveis. Os descendentes, em qualquer geração, serão ligeiramente diferentes em suas características (cor, tamanho, forma, etc) e muitas dessas características serão herdáveis.
Uma imagem com um grupo de 16 borboletas com asas em vários tons de azul e de vermelho. Um balão de texto que diz: Ei, vocês são vermelhas? Que legal! vem de uma das borboletas azuis. Um balão de texto que diz: Uau! Adorei essa cor de asa azul! vem de uma das borboletas vermelhas. O texto na parte inferior da imagem diz: Na verdade, as borboletas não falam! Desenho criado apenas para fins de ilustração bonitinha. Um rosto sorridente é mostrado ao lado do texto.
Baseado nessas observações, Darwin concluiu que:
Em uma população, alguns indivíduos vão herdar características que os ajudam a sobreviver e reproduzir (dadas as condições ambientais, como predadores e fonte de nutrientes disponíveis). Os indivíduos com essas características benéficas vão deixar uma prole maior na próxima geração quando comparados aos demais, já que tais características os tornam mais aptos a sobreviver e reproduzir.
Como as características úteis são herdáveis, e os organismos com essas vantagens deixam uma prole maior, elas se tornarão mais comuns (presentes em uma fração maior da população) na próxima geração.
Ao longo das gerações, a população se tornará adaptada ao seu ambiente (visto que indivíduos com características úteis a este ambiente têm, consistentemente, maior sucesso reprodutivo que os seus contemporâneos).
O modelo de evolução de Darwin por seleção natural permitiu-lhe explicar os padrões que ele havia visto durante suas viagens. Por exemplo, se as espécies de tentilhão de Galápagos compartilhassem um ancestral comum, faria sentido que elas se assemelhassem fortemente entre si (e aos tentilhões do continente, que provavelmente compartilhavam esse mesmo ancestral comum). Contudo, se grupos de tentilhões tivessem sido isolados em ilhas separadas por muitas gerações, cada grupo teria sido exposto a um ambiente diferente no qual diferentes características herdáveis poderiam ter sido favorecidas, tais como tamanhos e formatos diferentes de bico para a utilização de diferentes fontes de alimento. Esses fatores poderiam ter levado à formação de espécies distintas em cada ilha.
Exemplo: como a seleção natural funciona
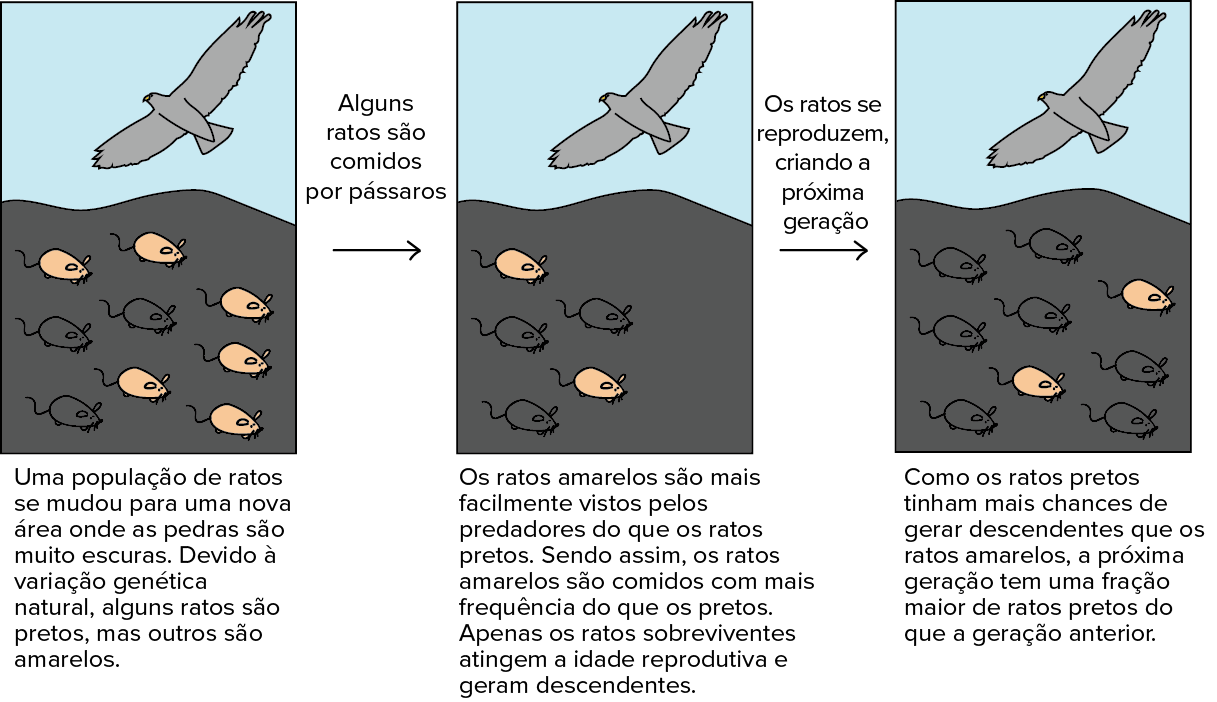
Para tornar a seleção natural mais concreta, consideremos um exemplo simplificado, hipotético. Nesse exemplo, um grupo de ratos com variação herdável da cor de pelo (preto vs. amarelo) acabou de se mudar para um nova área onde as rochas são pretas. Esse ambiente apresenta gaviões, que gostam de comer ratos e podem ver os amarelos mais facilmente que os pretos em contraste com a rocha preta.
Já que os gaviões podem ver e pegar os ratos amarelos mais facilmente, uma fração relativamente maior de ratos amarelos são comidos, enquanto uma fração muito menor de ratos pretos serão comidos. Se olharmos para a proporção entre ratos pretos e amarelos dentro do grupo sobrevivente ("não comidos"), ela será maior do que na população inicial.
A cor do pelo é uma característica herdável (uma que pode ser passada dos pais ao filho). Então, a fração aumentada de ratos pretos no grupo sobrevivente significa uma fração aumentada de filhotes pretos na próxima geração. Após várias gerações de seleção, a população pode ser composta quase inteiramente de ratos pretos. Essa mudança nas características herdáveis da população é um exemplo de evolução.
A teoria sintética da evolução
O neodarwinismo ou teoria sintética da evolução é uma teoria evolucionista que se baseia na teoria da evolução proposta por Darwin, porém adiciona conhecimentos científicos que possibilitaram explicar algumas lacunas apresentadas no darwinismo, por exemplo, como surge a variabilidade dentro de uma população. O neodarwinismo seria, portanto, basicamente uma reformulação da teoria de Darwin, incorporando, por exemplo, dados sobre genética.
A mutação amplia a variabilidade genética
No neodarwinismo, considera-se, além da seleção natural, que outros fatores evolutivos agem sobre as espécies, tais como mutação, recombinação, migração e deriva genética. As mutações e a recombinação tendem a aumentar a variação genética entre os indivíduos, e a seleção natural atua selecionando as variações que garantem maior chance de sobrevivência para o organismo.
Veja, a seguir, a definição dos fatores evolutivos considerados pelo neodarwinismo:
Mutação: é uma alteração no material genético que ocorre de maneira aleatória e repentina nos indivíduos. Caracteriza-se por ser afonte primária de variabilidade genética para as espécies. As mutações ocorrem de maneira espontânea, mas podem ser provocadas pela ação de agentes mutagênicos. Vale salientar que as mutações não ocorrem como forma de adaptar o indivíduo ao ambiente em que vive. Algumas mutações podem levar ao desenvolvimento de características pouco vantajosas e que, nesse caso, são eliminadas. As mutações positivas serão, por meio da seleção natural, mantidas na população.
Recombinação genética: bem como a mutação, também aumenta a variabilidade. Entretanto, na recombinação, temos um rearranjo dos genes já existentes em uma população; já na mutação, novos alelos são formados. A recombinação genética está presente no crossing-over(permuta de material genético entre cromátides não irmãs), que ocorre na prófase I da meiose, e na fusão de gametas (fecundação).
Deriva genética:é uma mudança nas frequências alélicas de uma geração para outra que ocorre devido a eventos aleatórios, como resultado do acaso. Os genes passados para as próximas gerações, nesse caso, não são aqueles que conferem maior sucesso adaptativo e sim aqueles que apresentaram mais “sorte”. A deriva genética, portanto, não atua para produzir adaptações. O efeito gargalo e o efeito fundador são dois casos de deriva genética. O efeito gargalo acontece quando fatores ambientais levam à repentina redução no tamanho da população, e o efeito fundador ocorre quando uma população pequena coloniza uma nova área. A deriva genética, diferentemente dos outros fatores estudados, atua diminuindo a variabilidade genética.
Bases genéticas da evolução
A mutação cria novos genes, e a recombinação os mistura com os genes já existentes, originando os indivíduos geneticamente variados de uma população. A seleção natural, por sua vez, favorece os portadores de determinados conjuntos gênicos adaptativos, que tendem a sobreviver e se reproduzir em maior escala que outros. Em função da atuação desses e de outros fatores evolutivos, a composição gênica das populações se modifica ao longo do tempo.
Mutações
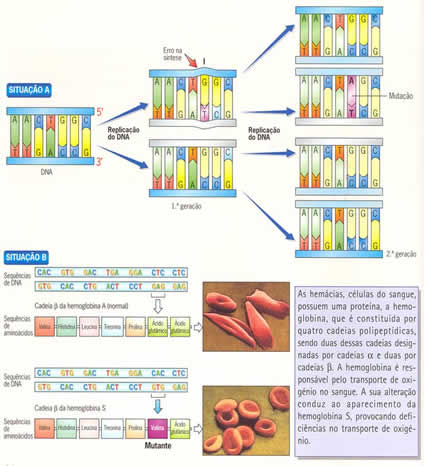
As mutações podem ser cromossômicas ou gênicas. As mutações cromossômicas podem ser alterações no número ou na forma dos cromossomos. As mutações gênicas originam-se de alterações na sequência de bases nitrogenadas de determinado gene durante a duplicação da molécula de DNA. Essa alteração pode ocorrer por perda, adição ou substituição de nucleotídeos, o que pode originar um gene capaz de codificar uma proteína diferente da que deveria ter sido codificada.
As mutações gênicas são consideradas as fontes primárias da variabilidade, pois aumentam o número de alelos disponíveis em um lócus, incrementando um conjunto gênico da população. Embora ocorram espontaneamente, podem ser provocados por agentes mutagênicos, como radiações e certas substâncias químicas (a droga ilegal LSD, por exemplo).
As mutações não ocorrem para adaptar o indivíduo ao ambiente. Elas ocorrem ao acaso e, por seleção natural, são mantidas quando adaptativas (seleção positiva) ou eliminadas em caso contrário (seleção negativa). Podem ocorrer em células somáticas ou em células germinativas; neste último caso as mutações são de fundamental importância para a evolução, pois são transmitidas aos descendentes.
O conceito de população mendeliana
Na linguagem diária, o termo “população” é muito empregado em contextos geográficos ou ecológicos para se referir à população de uma cidade, de um país ou de um determinado ambiente natural. Em genética, no entanto, usa-se uma definição mais específica:
População é um conjunto de indivíduos que se reproduzem sexuadamente, compartilhando um patrimônio gênico comum.
Essa definição foi proposta em 1950 por Theodosius Dobzhansky (1900-1975) e caracteriza o que se convencionou chamar população mendeliana, por aludir ao aspecto genético.
Em uma população mendeliana, com exceção dos gêmeos univitelinos, os indivíduos são diferentes um dos outros em uma serie de características. Cada indivíduo possui seu conjunto gênico particular, diferente do conjunto gênico de todos os demais membros da população.
Migração
A migração corresponde à entrada ou à saída de indivíduos em uma população. A entrada denomina-se imigração e a saída emigração.
Pelos processos migratórios é possível que genes novos sejam introduzidos em uma população. Assim, se indivíduos emigrarem de uma população para a outra da mesma espécie, poderão introduzir genes que não ocorriam na população para a qual imigraram, contribuindo para o aumento da variabilidade genotípica dessa população.
Por meio das migrações é estabelecido um fluxo gênico, que tende a diminuir as diferenças genéticas entre as populações da mesma espécie.
Frequências gênicas em populações
Considere uma par de alelos A e a. Em uma população hipotética de 10.000 indivíduos, suponha que 3.600 sejam homozigotos AA, 1600 sejam homozigotos aa e 4.800 sejam heterozigotos Aa. Nessa população há um total de 20.000 alelos do loco gênico considerado, uma vez que cada indivíduo apresenta um par deles. O número de alelos A é 12.000, pois os 3.600 indivíduos homozigotos AA apresentam um total de 7.200 alelos A, e os 4.800 heterozigotos Aa apresentam um total de 4800 alelos A (7.200 + 4.800 = 12.000).
AA
3.600
A = 3.600
12.000
20.000
A = 3.600
Aa
4.800
A = 4.800
a = 4.800
8.000
aa
1.600
a = 1.600
a = 1.600
A frequência de A é calculada dividindo-se o número total de alelos A (12.000) pelo número total de pares de alelos da população considerada (20.000). Portanto, nesse caso, a frequência de A é igual a 0,6 ou 60% (f(A) = 12.000/20.000 = 0,6).
A frequência do alelo a pode ser calculada da mesma maneira. Os 1.600 indivíduos homozigotos aa apresentam um total de 3.200 alelos a, e os 4800 heterozigotos Aa apresentam 4800 alelos a, totalizando 8.000 genes. Portanto, a frequência de a é igual a 0,40 ou 40% (f(a) = 8.000/20.000 = 0,4).
Alelos
Nº total de alelos
Nº total de alelos na população considerada
Frequência dos alelos na população
A
12.000
20.000
12.000/20.000 = 0,6
a
8.000
8.000/20.000 = 0,4
O segundo cálculo é desnecessário, uma vez que a soma das frequências dos alelos de um loco, em uma população, é sempre igual a 1. No caso:
f(A) + f(a) = 1 ou 100%
Utilizando no exemplo:
0,6 + 0,4 = 1
Consequentemente:
f(a) = 1-f(A)
Utilizando no exemplo:
f(a) = 1- 0,6
f(a) = 0,4
O princípio de Hardy-Weinberg
Quando o assunto é evolução e genética de populações, não podemos deixar de citar o princípio de Hardy-Weinberg, também conhecido por lei do equilíbrio de Hardy- Weinberg. Criado em 1908 pelo matemático Godfrey Hardy e pelo médico Wilhelm Weinberg, o princípio enfatiza que caso os fatores evolutivos, tais como seleção natural, mutação, migração e oscilação genética, não atuem sobre uma determinada população, as frequências gênicas e as proporções genotípicas permanecerão constantes. Isso quer dizer que se existem, por exemplo, os alelos B e b em uma população, eles não sofrem mudanças em suas taxas por um longo período de tempo. Essas taxas só seriam alteradas se ocorressem mecanismos evolutivos.
Para demonstrar o princípio de Hardy-Weinberg, uma população deve obedecer a algumas premissas. Primeiramente ela deve ser consideravelmente grande e apresentar o mesmo número de machos e fêmeas. Outro ponto importante é que todos os casais devem ser igualmente férteis e capazes de produzir o mesmo número de filhotes. Todos os cruzamentos devem ocorrer de forma aleatória. Por fim, não podem ocorrer mutações nessa população, ela não pode sofrer seleção natural e não pode ocorrer fluxo gênico. Percebe-se, portanto, que somente uma população teórica pode satisfazer esse princípio.
Podemos concluir que o princípio de Hardy-Weinberg pode ser usado como um indicativo de que determinada população sofreu evolução. Isso pode ser feito através da análise da frequência dos alelos. Caso a frequência mude, é sinal de que fatores evolutivos agiram ali.
Calcular a frequência de genes e genótipos de uma população em equilíbrio de Hardy-Weinberg é bastante simples. Suponhamos que existam o alelo B, que será representado por p, e um alelo b, que será representado por q, em uma população. A soma da frequência desses dois alelos deve ser igual a 100%, logo:
p+q=1
Continuando com essa população como exemplo, temos os seguintes genótipos: BB, Bb e bb. Para que um indivíduo seja BB, ele deve herdar um alelo B do pai e um alelo B da mãe, sendo assim, a frequência desse genótipo é p2. Da mesma maneira, a frequência de bb é q2. Já a frequência de Bb é 2pq, uma vez que o indivíduo pode receber o alelo B do pai ou da mãe e o alelo b da mesma forma. Temos, portanto, as seguintes frequências de genótipos:
F(BB)= p2
F(Bb)= 2pq
F(bb) = q2
Exemplo:
(Fuvest) Numa população de 100 pessoas, 36 são afetadas por uma doença genética condicionada por um par de alelos de herança autossômica recessiva.
a) Expresse, em frações decimais, a frequência dos genes dominantes e recessivos.
b) Quantos indivíduos são homozigotos?
c) Suponha que nessa população os cruzamentos ocorram ao acaso, deles resultando, em média, igual número de descendentes. Considere, também, que a característica em questão não altera o valor adaptativo dos indivíduos. Nessas condições, qual será a porcentagem esperada de indivíduos de fenótipo dominante na próxima geração?
Justifique suas respostas mostrando como chegou aos resultados numéricos.
Resolução:
a) Se uma população apresenta 100 pessoas e 36 são afetadas por uma doença autossômica recessiva, temos 36% de afetados, ou 0,36. 0,36 corresponde a q2. Então q é igual a 0,6. Como p+q=1, temos que p é igual a 0,4.
b) Os indivíduos homozigotos são os indivíduos com genótipo AA e aa. Temos, portanto:
F(AA)+ F(aa) = (0,6)2+ (0,4)2
F(AA)+ F(aa) = 0,36 +0,16 = 0,52 ou 52 indivíduos.
c) Os indivíduos que apresentam fenótipo dominante são aqueles com genótipo Aa e Aa. Obedecendo ao princípio de Hardy-Weinberg, a frequência dos alelos deve manter-se constante. Sendo assim, a frequência dos genótipos será a mesma na geração sequente. Temos, portanto:
F(AA)+ F(Aa) = p2+ 2pq
F(AA)+ F(Aa) = (0,4)2 + 2(0,4.0,6) = 0,64
Fatores que alteram o equilíbrio gênico
O equilíbrio de Hardy-Weinberg não ocorre em populações reais, pois elas são afetadas constantemente por fatores que influenciam as frequências alélicas e genotípicas, o que altera o seu equilíbrio. Alterações nas frequências alélicas e genotípicas ao longo do tempo levam à evolução.
As mutações permitem o surgimento de novos alelos nas populações.
Os fatores evolutivos que alteram o equilíbrio de Hardy-Weinberg são:
Migração: a chegada e a saída de indivíduos podem provocar mudanças nas frequências alélicas e genotípicas da população, pois genes estão sendo retirados e introduzidos. Assim, os indivíduos que estão chegando podem apresentar diferentes taxas de determinados genes e afetar as taxas que estavam presentes na população original.
Mutação: ocorrem ao acaso, sendo responsáveis pelo surgimento de novos alelos. Por meio delas, um alelo (A), por exemplo, pode dar origem a um novo alelo (a). Se esse alelo (a) apresentar uma maior viabilidade, ele será transmitido aos descendentes e se tornará mais frequente na população que o alelo (A).
Seleção natural: dentro de uma mesma população, os indivíduos apresentam variações nas suas características herdáveis. Indivíduos que apresentam características mais adequadas ao ambiente tendem a produzir uma maior prole do que aqueles sem essas características, que tendem a ser eliminados.
Oscilação gênica ou deriva genética: são alterações nas taxas de genes que ocorrem ao acaso, não por mutação ou pressão seletiva, em populações pequenas.
Deriva Gênica
A deriva genética corresponde a uma drástica alteração casual de ordem natural, atingindo a concentração genotípica de uma ou várias espécies, não preliminarmente envolvendo fatores de seleção natural, mas ocasionada por eventos repentinos.
Tal fenômeno é caracterizado pela ocorrência de catástrofes ecológicas, por exemplo: terremotos, tsunamis, tornados, inundações, queimadas, avalanches e outros processos, atingindo um grande contingente populacional.
Limitando, desda forma, o teor genético de um determinado grupo, restrito aos indivíduos prevalecentes. Cabendo a estes, integração a outra população, caso mantida uma adaptação, ou com o decorrer do tempo, a partir de um isolamento geográfico e posterior reprodutivo, constituição de uma nova espécie (princípio da espécie fundadora).
Nesta situação, portando uma baixa variabilidade, os indivíduos diferenciados irão passar por uma pressão de seleção mais significativa em relação à linhagem ascendente, que minimizava os feitos da seleção em razão do elevado número de indivíduos viventes.
Durante a evolução, uma hipótese referente à dizimação dos dinossauros, ocorrida por volta de 66 milhões de anos, estabelece não somente o fim destes animais, mas de variadas formas de vida (incluindo vegetais), em virtude de um imenso asteroide que teria atingido o planeta, levantando uma densa nuvem de poeira resultante da colisão, comprometendo os processos fotossintéticos (dependentes da energia solar), bem como a respiração e nutrição dos organismos.
Efeitos da deriva genética
A deriva genética pode ser explicada a partir de dois efeitos, o efeito gargalo e o efeito fundador. Entenda um pouco sobre cada um deles a seguir.
→ Efeito gargalo
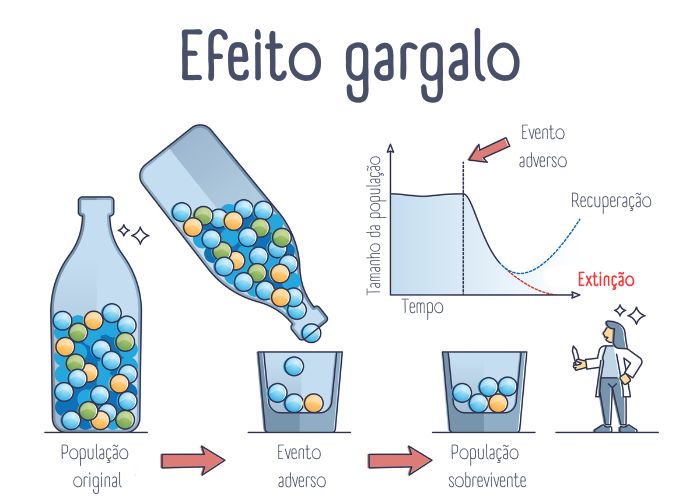
O efeito gargalo acontece quando uma população passa por uma redução significativa e abrupta em seu tamanho. Esse evento pode ocorrer devido a fatores naturais, como catástrofes ambientais, ou propositais, como caçadas intensivas e outros fatores que levam a uma diminuição drástica na quantidade de indivíduos da população.
Durante um efeito gargalo, apenas um pequeno número de indivíduos sobrevive e contribui para a próxima geração. Como resultado, a diversidade genética da população é reduzida, pois apenas um subconjunto limitado dos alelos presentes na população original é transmitido aos descendentes.
Utiliza-se “gargalo” porque o processo se assemelha ao estreitamento de um gargalo: as informações genéticas que sobrevivem ao evento são restritas.
Tendo como exemplo a imagem para entender como funciona o efeito gargalo, as bolinhas coloridas representam a diversidade genética de uma população, que é composta por alelos diferentes, cada qual representado por uma coloração. O gargalo da garrafa, por sua vez, representa um evento adverso que leva à diminuição abrupta da população e, consequentemente, acontece uma redução da diversidade genética dessa população. Observe que a população que sobrevive ao evento é composta por uma frequência de alelos diferente da população original. Após algum tempo, o tamanho da população se restabelece, porém existe uma diversidade genética menor do que a observada na população original.
Um exemplo do efeito gargalo acontece com os elefantes-marinhos-do-norte, que apresentam uma redução na variação genética, possivelmente devido a um evento de gargalo populacional causado pela interferência humana na década de 1890. A atividade de caça reduziu drasticamente suas populações para aproximadamente 20 indivíduos no final do século XIX. Embora essas populações tenham se recuperado desde então, ultrapassando os 30.000 indivíduos, os efeitos do gargalo persistem em seus genes. Em comparação com uma população de elefantes-marinhos-do-sul, que sofreu uma caça menos intensiva, os elefantes-marinhos-do-norte exibem menor variação genética.
→ Efeito fundador
O efeito fundador ocorre quando um pequeno grupo de indivíduos se separa de uma população maior e inicia uma nova população em uma área geográficaseparada. Esses fundadores, por vezes, representam apenas uma fração da diversidade genética presente na população original de onde se originaram. Como resultado, a nova população fundada pode ter uma composição genética mais limitada e menos diversificada do que a população original.
Um exemplo do efeito fundador é o que ocorre na população africana de descendentes de colonos holandeses no sul da África. Essa população tem sua origem principalmente em um pequeno grupo de colonos holandeses iniciais. Atualmente, essa população apresenta uma alta frequência do gene associado à doença de Huntington, pois os colonos holandeses originais coincidentemente carregavam genes com uma frequência notavelmente elevada para essa condição.
Consequências da deriva genética
A deriva genética gera consequências na variabilidade genética e na fixação ou redução dos alelos presentes em uma população. Algumas delas são:
Fixação de alelos: um alelo específico se torna presente em 100% da população. Isso ocorre por acaso, e alelos fixados podem não necessariamente conferir vantagens adaptativas.
Perda de alelos: o oposto ocorre em relação à fixação, isto é, um alelo específico é eliminado de uma população ao longo do tempo, o que implica o seu desaparecimento nessa população.
Perda da variabilidade genética: a fixação de apenas alguns alelos e o desaparecimento de outros resultam da diminuição da variabilidade genética, o que pode tornar a população mais suscetível a mudanças que ocorram no ambiente, como mudanças climáticas ou alteração dos recursos disponíveis.
Diferenças entre mutação e deriva genética
A deriva genética é um evento diferente da mutação porque gera variações na frequência genética de uma população a partir de mecanismos diferentes daqueles que ocorrem na mutação.
Mutação: é uma alteração no material genético de um organismo, mais especificamente no seu DNA. Isso pode ocorrer de diversas maneiras, como substituições, deleções ou inserções de nucleotídeos, que acontecem naturalmente e ao acaso durante a replicação do DNA ou em resposta a fatores ambientais, radiação ou agentes químicos. As mutações introduzem variações genéticas que podem ser hereditárias e que contribuem para a diversidade genética nas populações.
Deriva genética: é um processo evolutivo que não introduz novas variantes genéticas, mas atua na distribuição de alelos existentes em uma população devido a flutuações aleatórias nas frequências alélicas ao longo do tempo. Enquanto as mutações representam alterações no material genético, a deriva genética se relaciona com a variação aleatória da composição genética de uma população.
A reprodução é um processo biológico que se refere à capacidade que os seres vivos apresentam de gerar descendentes. As duas grandes categorias que agrupam os tipos de reprodução existentes são: reprodução sexuada e reprodução assexuada . A reprodução é fundamental para a preservação e continuidade das espécies, pois através dela as informações genéticas dos organismos preexistentes são transmitidas aos organismos criados. As principais diferenças entre os métodos de reprodução são: A reprodução assexuada é geralmente mais simples e mais rápida que a reprodução sexuada. A reprodução assexuada envolve apenas um membro da espécie, enquanto a reprodução sexuada é realizada por dois progenitores. A reprodução assexuada forma organismos geneticamente idênticos, enquanto na reprodução sexuada ocorre recombinação gênica. A reprodução sexuada produz variabilidade genética, ao contrário da reprodução assexuada que produz clones. Reprodução assexuada Na reprod...
Sobre a Biotecnologia A biotecnologia utiliza células vivas para desenvolver ou manipular produtos com fins específicos, como, por exemplo, os alimentos transgênicos. Assim sendo, a biotecnologia está vinculada com a engenharia genética e surgiu como disciplina no começo do século XX na indústria alimentar , à qual, posteriormente, se somaram outros setores como a medicina ou o meio ambiente. Atualmente, os cinco ramos nos quais se divide a biotecnologia moderna — humana, ambiental, industrial, animal e vegetal — nos ajudam a combater a fome e as doenças, produzir de forma mais segura, limpa e eficiente, reduzir nossa pegada ecológica e poupar energia. O aumento do investimento e do emprego nesse setor tem sido exponencial nos últimos anos, o que reflete seu crescente impacto econômico e sua importância para o desenvolvimento sustentável e a inovação tecnológica em todo o mundo. Usos e aplicações da biotecnologia As inovações biotecnológicas já fazem...
A primeira Lei de Mendel diz que as características dos indivíduos são condicionadas por pares de fatores (genes) e que estes fatores se separam (segregam) durante a formação dos gametas. Quando estudamos esta lei da genética, geralmente nos remetemos à exemplos de dominância completa, onde um gene pode inibir a ação do outro quando ocorrer heterozigose. Porém, a primeira Lei de Mendel pode também se aplicar a outros mecanismos de herança, como a codominância e também a dominância incompleta. Dominância incompleta No tipo mais estudado de herança genética, a dominância completa, um dos alelos envolvidos na produção de uma característica domina o outro quando em heterozigose. Isso quer dizer que o alelo dominante vai impedir o outro gene de produzir sua característica. Neste tipo de herança, os descendentes sempre terão o fenótipo de um dos progenitores. Mas, como já disse anteriormente, nem todos os mecanismos genéticos seguem essa regra. Em algu...















Comentários
Postar um comentário